Recently, Lee et al.designed chemically modified peptidic derivates of a Taspase1 cleavage substrate. Although some of these compounds displayed mild SCH772984 inhibitory activity using in vitro Taspase1 assays, these peptide-based inhibitors have not shown efficacy in living cells, in contrast to our low molecular weight inhibitors. Although natural products appear to interrogate a different area of chemical space than synthetic compounds, the tested lipophilic fungal extracts showed no inhibitory activity. Failure may be due to the fact that albeit such extracts contain a mixture of many different substances, the concentration of potentially active ingredients may be too low or outweighed by toxic effects of other components. Also, the numbers of samples which have to be screened in unfocussed natural product libraries are usually high, and hit rates are mostly below 0.01%. Hence, as future strategies to identify potent Taspase1 inhibitors we suggest to focus on a rational synthesis of derivates based on the structures of our primary hits combined with HTS of large natural/synthetic compound libraries. Automatic thresholding using the Isodata algorithm was used to convert the image to a binary mask that included all fluorescence data above background. The Hoechst 33342 stainingmask was used to define the nuclear ROI. Subsequently, the Hoechst 33342 mask was subtracted from the GFP maskto create a staining mask defining the cytoplasmic ROI. Scans were performed sequentially with settings to give sub-saturating fluorescence intensity, and a minimum of 400 valid objects per well was recorded. Acid sphingomyelinaseis a lysosomal glycoprotein that catalyses the hydrolysis of sphingomyelin into ceramide and phosphorylcholine. Fusion of secretory lysosomes with the cell surface and translocation of lysosomal ASM onto the outer leaflet of the cell membrane plays an important role during stress response. CD95 ligands and cytokines such as tumor necrosis factor-a, interleukin-1 and interferon-c but also other stimuli including oxidative stress, reactive oxygen and nitrogen species, ionizing radiation, UV-C radiation, heat shock and other agents of stress, injury or infections by HIV or bacteria have been shown to stimulate ceramide production, assumed to be in part due to increased ASM activity. Ceramide, in turn, leads to membrane reorganization and downstream signalling that results in cell activation, very often cell stress or apoptosis. In addition to ASM, at least three other sphingomyelinases have been described in mammalian cells that vary in their pH optimum and cofactor dependency. Although these enzymes and an existing de novo synthesis pathway are alternative mechanisms for ceramide generation, activation of ASM itself 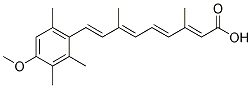 has been proven to be critical for some cellular responses, such as WZ8040 apoptosis induced by reactive oxygen and nitrogen species, chemotherapy drugs such as cisplatin, bacteria, radiationand CD95. Furthermore, in contrast to other sphingomyelinases, ASM activity is tightly regulated. Ceramide is further metabolized to sphingosine and sphingosine-1-phosphate by acid ceramidaseand sphingosine kinases. While the biological function of sphingosine is largely unknown, sphingosine-1-phosphate has been shown to be involved in cellular differentiation, proliferation and cell migration. This dynamic balance between ceramide and sphingosine-1-phosphate is referred to as the “ceramide/sphingosine-1phosphate rheostat”, maintaining the balance between growth and cell death. ASM is best known for its involvement in Niemann-Pick disease, a lysosomal storage disease due to an inherited enzyme deficiency.
has been proven to be critical for some cellular responses, such as WZ8040 apoptosis induced by reactive oxygen and nitrogen species, chemotherapy drugs such as cisplatin, bacteria, radiationand CD95. Furthermore, in contrast to other sphingomyelinases, ASM activity is tightly regulated. Ceramide is further metabolized to sphingosine and sphingosine-1-phosphate by acid ceramidaseand sphingosine kinases. While the biological function of sphingosine is largely unknown, sphingosine-1-phosphate has been shown to be involved in cellular differentiation, proliferation and cell migration. This dynamic balance between ceramide and sphingosine-1-phosphate is referred to as the “ceramide/sphingosine-1phosphate rheostat”, maintaining the balance between growth and cell death. ASM is best known for its involvement in Niemann-Pick disease, a lysosomal storage disease due to an inherited enzyme deficiency.
Pathological reduction of ASM activity may be caused by mutations in the ASM gene itself
Leave a reply