Stimulation with GM-CSF brings the modifying enzyme and the bait in close proximity, while interaction of the modified bait with a MAPPIT prey is detected as in the regular MAPPIT setup. In our currently presented method, we used degenerated PCR via Mutazyme II or random mutagenesis, as it allows a low mutation frequency necessary to obtain single point mutations. Catharanthine sulfate Moreover, it allows all types of transitions and transversions and shows a reasonably balanced distribution of mutations among the different codons. This reduces the risk of an unbalanced distribution of mutations along the sequence, which could bias our analysis.The Mutazyme II allows a good control of the number of mutations by varying the number of PCR cycles and the amount of input DNA. However, the exact conditions probably differ for different proteins and need to be optimized before the 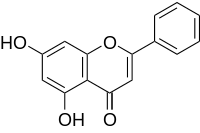 MAPPIT analysis. Many different methods for the Chlorhexidine hydrochloride introduction of random mutations have been described and most methods are probably compatible with the method presented in this paper. For example, the introduction of mutations via degenerated primers or gene synthesis can allow the introduction of random mutations at specific sites, and increase the number of single amino acid substitutions. Interestingly, scanning mutagenesis methods via mu transposase variants allow the random integration, deletion or replacements of single or multiple codons by one or more specific codons, allowing for example a random replacement of amino acids by a specific amino acid type. For the development of the random mutagenesis strategy based on MAPPIT, dimerization of Apobec3G and its interaction with Vif and Gag were used as targets. In the absence of a structure of Vif or of the full-length Apobec3G protein, several molecular aspects of the interplay between Apobec3G and Vif remain unclear. In a previous study, the importance of a predicted headto-head interface in the N-terminal domain was tested via site directed mutagenesis and MAPPIT. The study confirmed the importance of this interface for Apobec3G-Apobec3G interaction, but also demonstrated that the interface is required for Vif binding. The current study shows that the head-to-head interface of the N-terminal domain is also important for binding to Gag. This correlates well with previous studies that showed the importance of the N-terminal Apobec3G domain for the high affinity interaction with Vif and the RNA-mediated interaction with Gag. The C-terminal domain of Apobec3G is the catalytic deaminase domain and is the target of polyubiquitination via Vif. MAPPIT analysis indicates that the C-terminal domain of Apobec3G is also important for interaction with Apobec3G, Vif and Gag. We therefore tried to identify which regions in the N- and C-terminal domain of Apobec3G are involved in the different interactions via random mutagenesis. A surprising outcome of the random mutagenesis MAPPIT analysis is that no single amino acid substitutions were found that specifically affect only one of the three tested interactions of Apobec3G. A possible explanation for that is that specific binding sites were missed because the coverage of our mutagenesis analysis was too low. However, several arguments argue against this. In the site-directed mutagenesis analysis of the N-terminal head-to-head interface, we found that mutations of 17 residues affected the interaction with Vif, Apobec3G or Gag by more than 50%. The random mutagenesis screen detected 12 of these residues, suggesting 70% coverage. This coverage goes together with some redundancy: 47 of the 114 mutated residues discovered in the screen are found multiple times, often with different substitutions. In the ongoing random mutagenesis analyses of Mal and RNF41, we do find multiple mutations that specifically affect one of the interactions without affecting the other interactions.
MAPPIT analysis. Many different methods for the Chlorhexidine hydrochloride introduction of random mutations have been described and most methods are probably compatible with the method presented in this paper. For example, the introduction of mutations via degenerated primers or gene synthesis can allow the introduction of random mutations at specific sites, and increase the number of single amino acid substitutions. Interestingly, scanning mutagenesis methods via mu transposase variants allow the random integration, deletion or replacements of single or multiple codons by one or more specific codons, allowing for example a random replacement of amino acids by a specific amino acid type. For the development of the random mutagenesis strategy based on MAPPIT, dimerization of Apobec3G and its interaction with Vif and Gag were used as targets. In the absence of a structure of Vif or of the full-length Apobec3G protein, several molecular aspects of the interplay between Apobec3G and Vif remain unclear. In a previous study, the importance of a predicted headto-head interface in the N-terminal domain was tested via site directed mutagenesis and MAPPIT. The study confirmed the importance of this interface for Apobec3G-Apobec3G interaction, but also demonstrated that the interface is required for Vif binding. The current study shows that the head-to-head interface of the N-terminal domain is also important for binding to Gag. This correlates well with previous studies that showed the importance of the N-terminal Apobec3G domain for the high affinity interaction with Vif and the RNA-mediated interaction with Gag. The C-terminal domain of Apobec3G is the catalytic deaminase domain and is the target of polyubiquitination via Vif. MAPPIT analysis indicates that the C-terminal domain of Apobec3G is also important for interaction with Apobec3G, Vif and Gag. We therefore tried to identify which regions in the N- and C-terminal domain of Apobec3G are involved in the different interactions via random mutagenesis. A surprising outcome of the random mutagenesis MAPPIT analysis is that no single amino acid substitutions were found that specifically affect only one of the three tested interactions of Apobec3G. A possible explanation for that is that specific binding sites were missed because the coverage of our mutagenesis analysis was too low. However, several arguments argue against this. In the site-directed mutagenesis analysis of the N-terminal head-to-head interface, we found that mutations of 17 residues affected the interaction with Vif, Apobec3G or Gag by more than 50%. The random mutagenesis screen detected 12 of these residues, suggesting 70% coverage. This coverage goes together with some redundancy: 47 of the 114 mutated residues discovered in the screen are found multiple times, often with different substitutions. In the ongoing random mutagenesis analyses of Mal and RNF41, we do find multiple mutations that specifically affect one of the interactions without affecting the other interactions.
Coupled to two separate receptor chains with the extracellular part of the chain of the GM-CSF receptor
Leave a reply