This observation is consistent with the results of Anderson et al., who found that Plasmodiuminfected mosquitoes with sporozoites in their salivary glands were less likely to give up their Albaspidin-AA attempt to feed than uninfected mosquitoes. This change in insect behavior, with a larger number of probing attempts, may be accounted 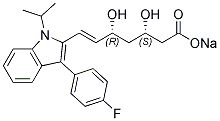 for by changes in the expression of salivary gland components or a modulation of the nervous system by the parasite. However, we observed no difference in probing time between Plasmodium berghei-infected and uninfected A. gambiae in this study. This observation is consistent with the findings of Pumpuni et al., who found no difference in total feeding time between Plasmodium yoelli-infected and uninfected Anopheles stephensi. The ability of mammals to produce antibodies against mosquito saliva antigens is well established. We therefore investigated whether the presence of antibodies in the blood of animals interfered with the blood meal. We found that immunized mice were more attractive to mosquitoes than non immunized mice and that the mosquitoes probed the immunized mice more rapidly. For feeding the first step in the behavioural sequence is to sample the substrate and determine if the food is suitable. This happens if the appropriate feeding stimulus, a phagostimulant, is present. Presence or absence of the phagostimulant results in either feeding or not, and in its absence the insect moves on continuing to look for food. showed an enhanced feeding success of Aedes aegypti mosquitoes on parasitemic hosts. In our case, it seems that the presence of anti-saliva antibodies could act indirectly as a phagostimulant. In mosquito sensitized against saliva, we can expect a modification of the skin by the interaction of saliva with the innate immune system, resulting in a pro-inflammatory host response causing a vasodilatation of blood vessels. In mice passively immunized, we propose that antibodies that have been passively transferred may modify the site of bite as shown by Interestingly, the biting mosquitoes tended to carry out capillary feeding in significantly larger blood 4-(Benzyloxy)phenol vessels in immunized than in naive mice. This observation may be explained because of the vasodilatation due to the pro-inflammatory response induced in saliva-sensitized mice. It could also be explained by the presence of anti-saliva antibodies in smaller vessels that may completely inhibit blood-feeding. This observation has potential implications, because the concentration of parasite stages infectious for mosquitoes may vary with vessel size. The concentration of gametocytes is higher in capillaries, whereas the percentage of old trophozoites is higher in larger vessels. By contrast, microfilarial density in small peripheral blood vessels has been shown to be lower than that in large blood vessels. A white area developed in direct contact with the proboscis of mosquitoes feeding on salivary-sensitized animals or mice injected with anti-saliva antibodies. Precipitating antibodies have been found in the blood of guinea pigs or rabbits bitten by Aedes aegypti. The white area may therefore correspond to an immune reaction to saliva delivered to the bloodstream during blood feeding. This assumes that mosquitoes salivate during blood feeding, resulting in the exposure of saliva antigens to blood components. This hypothesis is entirely tenable, because Griffiths and Gordon observed mosquito salivation within a blood vessel during blood feeding and Kebaier et al. reported an apparent precipitant reaction at the distal end of the mosquito proboscis.
for by changes in the expression of salivary gland components or a modulation of the nervous system by the parasite. However, we observed no difference in probing time between Plasmodium berghei-infected and uninfected A. gambiae in this study. This observation is consistent with the findings of Pumpuni et al., who found no difference in total feeding time between Plasmodium yoelli-infected and uninfected Anopheles stephensi. The ability of mammals to produce antibodies against mosquito saliva antigens is well established. We therefore investigated whether the presence of antibodies in the blood of animals interfered with the blood meal. We found that immunized mice were more attractive to mosquitoes than non immunized mice and that the mosquitoes probed the immunized mice more rapidly. For feeding the first step in the behavioural sequence is to sample the substrate and determine if the food is suitable. This happens if the appropriate feeding stimulus, a phagostimulant, is present. Presence or absence of the phagostimulant results in either feeding or not, and in its absence the insect moves on continuing to look for food. showed an enhanced feeding success of Aedes aegypti mosquitoes on parasitemic hosts. In our case, it seems that the presence of anti-saliva antibodies could act indirectly as a phagostimulant. In mosquito sensitized against saliva, we can expect a modification of the skin by the interaction of saliva with the innate immune system, resulting in a pro-inflammatory host response causing a vasodilatation of blood vessels. In mice passively immunized, we propose that antibodies that have been passively transferred may modify the site of bite as shown by Interestingly, the biting mosquitoes tended to carry out capillary feeding in significantly larger blood 4-(Benzyloxy)phenol vessels in immunized than in naive mice. This observation may be explained because of the vasodilatation due to the pro-inflammatory response induced in saliva-sensitized mice. It could also be explained by the presence of anti-saliva antibodies in smaller vessels that may completely inhibit blood-feeding. This observation has potential implications, because the concentration of parasite stages infectious for mosquitoes may vary with vessel size. The concentration of gametocytes is higher in capillaries, whereas the percentage of old trophozoites is higher in larger vessels. By contrast, microfilarial density in small peripheral blood vessels has been shown to be lower than that in large blood vessels. A white area developed in direct contact with the proboscis of mosquitoes feeding on salivary-sensitized animals or mice injected with anti-saliva antibodies. Precipitating antibodies have been found in the blood of guinea pigs or rabbits bitten by Aedes aegypti. The white area may therefore correspond to an immune reaction to saliva delivered to the bloodstream during blood feeding. This assumes that mosquitoes salivate during blood feeding, resulting in the exposure of saliva antigens to blood components. This hypothesis is entirely tenable, because Griffiths and Gordon observed mosquito salivation within a blood vessel during blood feeding and Kebaier et al. reported an apparent precipitant reaction at the distal end of the mosquito proboscis.
During intravital microscopy of infected mosquitoes feeding on mice previously passively immunized with antibodies
Leave a reply
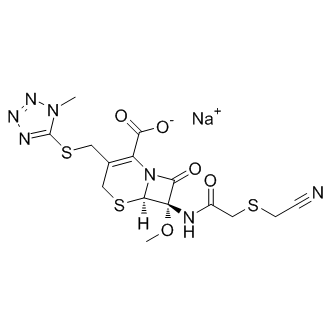 effects on many cell types, including proliferation, differentiation, and survival. IL-3 is a cytokine that induces growth and differentiation of hematopoietic stem cells and a variety of cell types originating in the bone marrow. Recent studies have demonstrated the important role of IL3 in the central nervous system. It is expressed in the hippocampus and cortices of normal mouse brain, and it stimulates the growth and proliferation of microglial cells. Studies also found that IL3 facilitates the survival of sensory neurons significantly and stimulates the formation of the neural network. In addition, IL-3 has been found to be able to promote the process extension of cultured cholinergic and prevent delayed neuronal death in the hippocampus. In fact, rat interleukin 3 receptor bsubunit was cloned from cultured microglia, and disruption of IL3 production in brain led to neurologic dysfunction. All of these studies strongly suggest that IL3 is a pivotal protective factor for CNS. More importantly, Chen et al. recently reported that IL3 was significantly associated with brain disease such as schizophrenia in three independent Irish samples.
effects on many cell types, including proliferation, differentiation, and survival. IL-3 is a cytokine that induces growth and differentiation of hematopoietic stem cells and a variety of cell types originating in the bone marrow. Recent studies have demonstrated the important role of IL3 in the central nervous system. It is expressed in the hippocampus and cortices of normal mouse brain, and it stimulates the growth and proliferation of microglial cells. Studies also found that IL3 facilitates the survival of sensory neurons significantly and stimulates the formation of the neural network. In addition, IL-3 has been found to be able to promote the process extension of cultured cholinergic and prevent delayed neuronal death in the hippocampus. In fact, rat interleukin 3 receptor bsubunit was cloned from cultured microglia, and disruption of IL3 production in brain led to neurologic dysfunction. All of these studies strongly suggest that IL3 is a pivotal protective factor for CNS. More importantly, Chen et al. recently reported that IL3 was significantly associated with brain disease such as schizophrenia in three independent Irish samples.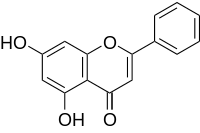 MAPPIT analysis. Many different methods for the
MAPPIT analysis. Many different methods for the 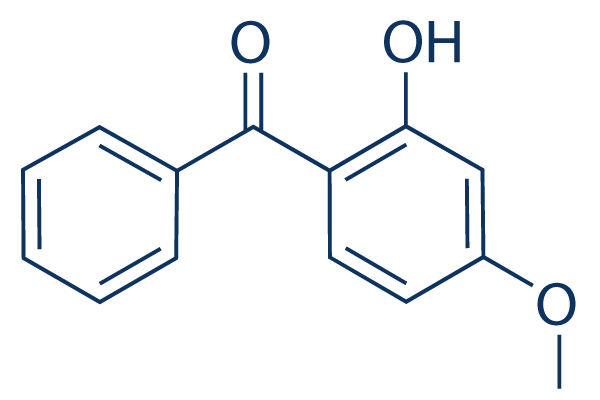 findings indicate that the expression of AnxA2 correlates with the aggressiveness of breast cancer and
findings indicate that the expression of AnxA2 correlates with the aggressiveness of breast cancer and  means of immune evasion, it is likely that functional antibodies to multiple invasion ligands would be needed to provide effective immunity. Demonstrating the functional significance of PfRh4 antibodies, affinity-purified human antibodies against the PfRh4.9 protein, which contains the erythrocyte-binding region, were potent inhibitors of SA-independent invasion, which requires the PfRh4 ligand.
means of immune evasion, it is likely that functional antibodies to multiple invasion ligands would be needed to provide effective immunity. Demonstrating the functional significance of PfRh4 antibodies, affinity-purified human antibodies against the PfRh4.9 protein, which contains the erythrocyte-binding region, were potent inhibitors of SA-independent invasion, which requires the PfRh4 ligand.